
The Physical Law of Evolutionary Stability – a Mechanistic Basis of any Ecological Structure
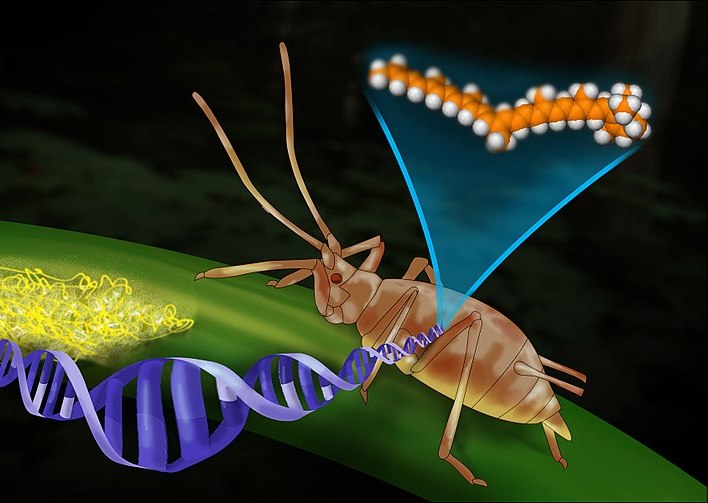
Living matter is regulated not only by the natural orders that also operate at the levels of non-living matter, but also by a structure of its own physical laws of nature. These can be recognized and proven according to the standards of empirical physics. Their central parameter consists of uncontrollably high degrees of complexity within all genomes and organisms and, consequently, also within ecosystems. A fundamental law in the uncontrollable complexities of ecological structures is the Law of Evolutionary Stability (LES), as described below. Its existence can be proven in several ways. It would have been of great importance for valid reasons to include it in the center of the natural sciences within the framework of empirical physics—because the agricultural methods of civilization run diametrically counter to this order and therefore cannot function sustainably.
According to the The Law of Evolutionary Stability, the uncontrollable high degree of complexity within an ecological structure makes it impossible for one species to sustainably determine any characteristics of the subsequent generations of another species for its own benefit. The adjective “sustainable” is important in this statement, as it refers to another concrete physical parameter that will become clear in a moment.
Preliminary remark
The following analysis of the physical Law of Evolutionary Stability (LES) and the demonstration of the unsustainability and resulting dysfunctionality of agricultural methods is not about disputing the food supply of eight billion people. On the contrary, only this open confrontation can offer a chance to prevent humanity from ending up in a final slum within a dying ecosystem. What such a global finale with escalating famines and misery would look like can already be seen in some regions. According to the criteria of true reason, the physical connections discussed here should be treated with the highest priority in the academic natural sciences. Open confrontation is unpleasant, but it is an indispensable prerequisite for finding ways to defuse the situation – which are still theoretically possible. Laws of nature can never be overcome. Continuing to turn a blind eye to the problem would mean abandoning the human children to a grim fate.
Even among viruses and microorganisms, there are many theoretical possibilities for the development of strategies in which one species influences the characteristics of future generations of another species in order to create its own primary benefit. This would then correspond in principle to the pattern that humans today follow through artificial selection and genetic engineering. Manipulative interference in genomes is virtually an everyday occurrence between microbes. However, the effects that are advantageous to the manipulating side always remain at the level of the individuals affected on the manipulated side. Among the several million empirical descriptions of parasitic interactions between different species, there seem to be no cases to be found in which these manipulations extend beyond the current generation of the attacked side and affect subsequent generations [4]. By logical reasoning, there can only be one possible reason for the absence of such strategies: namely, that they do not work sustainably and therefore could never have become established in the course of evolution.
The central mechanism of the Law of Evolutionary Stability is that all forms of life remain stable across generations in the uncontrollably complex ecological structure only if their characteristics are primarily aligned with their own benefit in natural selection. As soon as this is prevented by another life form for its own benefit through manipulative control of evolutionary development, the sustainability parameter automatically falls below zero and stability declines. It does not matter how the manipulation takes place. The affected hereditary lines of the manipulated side and, consequently, a manipulator specializing in them inevitably move into an evolutionary dead end.
The civilizational agricultural methodology of artificial selective breeding for the primary benefit of humans thus corresponded to the pattern of such a dead end from the outset. That it appeared to work for several millennia was due to the fact that very large quantities of original genetic resources from numerous plants and animals were virtually burned up, which took some time due to the enormous volume involved. This “burning” led to a short-term sharp increase in the amount of food available and, as a causal consequence, to a population explosion. Now, however, more than eight billion people exist on an increasingly unstable basis of genetically highly degenerated and unviable organisms.
It should be noted here that the Law of Evolutionary Stability is not a discovery of the authors of this article. Rather, the orders in question have been described before, albeit without a formal definition. The researcher Charles Darwin placed them at the center of his theory as a key insight. He wrote several times that his theory would be destroyed if even a single example were discovered anywhere in nature in which any characteristic of any species had arisen for the exclusive benefit of another species. The following is a relevant passage from his magnum opus “On the Origin of Species” from 1859:
„Natural selection cannot possibly produce any modification in any one species exclusively for the good of another species; though throughout nature one species incessantly takes advantage of, and profits by, the structure of another. (…) If it could be proved that any part of the structure of any one species had been formed for the exclusive good of another species, it would annihilate my theory, for such could not have been produced through natural selection. Although many statements may be found in works on natural history to this effect, I cannot find even one which seems to me of any weight.„ [1] Charles Darwin, 1859. (The original passage is linked to Darwin Online in the references.)
So Darwin was so sure that he even tied his entire theory inextricably to this one point. If someone had ever proven that a species in nature had developed a trait that was only useful to another species, then essentially the entire work of the famous naturalist would have collapsed.
But in fact, he has remained proven right to this day, thereby establishing empirical evidence by process of elimination when viewed retrospectively from the present. Although there are still occasional claims of alleged counterevidence, these always dissolve as soon as they are examined more closely. Sometimes such misrepresentations even come from academic life scientists, which, in addition to the mechanisms of collective repression, may be due, among other things, to the severe underdevelopment of the field with regard to the larger ecological context or to influences from the agricultural industry. Examples of this can be found in the German ZEIS Magazine.
Also in his magnum opus, Darwin stated with great clarity that the artificial selection of plants and animals by humans, i.e., the basis of civilized agricultural methodology, runs diametrically counter to the regularity described in the first quote, because it produces characteristics in other living beings that are only useful to humans. Here is a second quote, also from his magnum opus:
„One of the most remarkable features in our domesticated races is that we see in them adaptation, not indeed to the animal’s or plant’s own good, but to man’s use or fancy.„ [2]
Darwin recognized the objective connections solely through the observation of plants and animals. In his day, microbiology and genetics were virtually non-existent. This clearly shows that uncontrollable complexity is visible on the surface of ecological events, while beneath the surface it extends all the way down to the smallest and invisible structures.
The mechanical background of the physical Law of Evolutionary Stability will now be examined in more detail. It can be pointed out in advance that, despite millions of empirical descriptions of often extremely complex symbiotic, parasitic, and other relationships between viruses, microorganisms, fungi, plants, and animals, not a single example of even the smallest characteristic in any species has been proven to have developed for the exclusive benefit of another species.
When considering this, the multi-stage history of the development of the respective characteristics must always be taken into account. For example, if an insect benefits from the flower of a plant because it provides nutritious nectar and is also conspicuous in color, then the development of these characteristics was always based on the primary benefit to the plant, because the insect promoted its fertilization and reproduction. In natural selection, the more colorfully conspicuous variants of plants consequently had a relative advantage and passed on their characteristics to subsequent generations more frequently than their paler counterparts.
It is also important to understand that the direction we have taken in our agricultural methods, which goes against the Law of Evolutionary Stability, was not a case of our abstract thinking abilities or other characteristics of our intelligence outsmarting the “primitive” orders of nature. Here, too, the opposite is true: the processes within ecological structures are many times more complex than all our actions in the course of agricultural methodology, including the most modern methods of genetic engineering. For example, the CRISPR-Cas process is based on mechanisms that have developed convergently in many species of bacteria over at least hundreds of millions, but probably even several billion, years. However, they only use them at the individual level to eliminate parasitic phages.
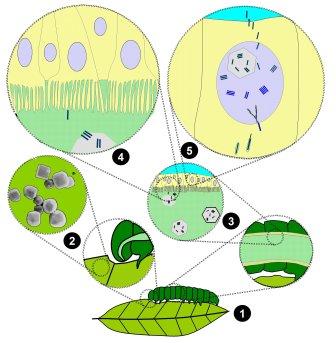
A vivid example that helps us understand that the absence of ‘breeding’ in nature has nothing to do with a lack of theoretical possibility, complexity or specificity is the reproductive strategy of viruses from the Baculoviridae family, which has probably existed for at least 300 million years: They manipulate caterpillars in an extremely precise manner so that those first crawl up to the highest tops of trees, contrary to their healthy behaviour. Once there, the virus takes control of the caterpillars‘ feeding instincts and cut off the natural feed stop. As a result, the infected larvae swell abnormally, while their body fluids become saturated with the virus. Eventually, they burst. The dripping droplets contaminate the leaves in the lower regions of the plant and thus the food of the caterpillars that are still healthy there. Before they burst, the infected caterpillars attach themselves to the treetops with a silk thread to ensure that they do not fall down before the droplets fall. This multi-stage parasitic strategy, which has evolved over millions of years of evolutionary selection, is precisely encoded in the genetic material of the viruses and includes, among other things, highly targeted interventions in the nervous system, metabolism and hormone balance of the caterpillars. The overall complexity and precision far exceed those of artificial selective breeding and genetic engineering practised by humans in agriculture. If targeted ‘breeding’ in the sense of influencing evolution between different forms of life to achieve sustainable advantages for the manipulator were to work, then a wide variety of such strategies would have had to have developed at the level of viruses and microorganisms during the billions of years of evolutionary selection. But not a single example, however minor, has ever been found. All parasites only ever manipulate the current generations of their hosts [4].
Indirect mathematical proof of uncontrollably high complexities as the basis of the Law of Evolutionary Stability
An essential mechanical basis of the Law of Evolutionary Stability can be indirectly proven through mathematics. It consists of the fact that the storage capacities of the genetic information of every life form, down to the relatively “simplest” microorganisms and even viruses, is extremely high, tending towards infinity, and that over a process of evolution lasting hundreds of millions or even several billion years, they have (had to) filled themselves with information in order to form an organization capable of surviving as sustainably as possible in a practically infinitely complex and constantly changing environment. The fact that the following “mathematical proof” is only indirect proof will soon become apparent as irrelevant. This is because the results of the calculations are so incredibly extreme that it would be practically irrelevant if, given the possibility of performing direct calculations, only a thousandth or a millionth of these results were to emerge.
To begin to understand these incomprehensible complexities, it is best to first look at the mechanical basics of the genetic memory of all living beings. At first glance, these function in a surprisingly simple way. In all forms of life, this occurs in a variable sequence of identical molecules of the four nucleotide bases adenine, thymine, guanine, and cytosine, abbreviated as A, T, G, and C (note: RNA viruses are excluded for sake of simplicity). The molecules are held in pairs (“base pairs”) by various auxiliary substances and are arranged in long strands. The entire mixture of nucleotides and auxiliary substances is called “deoxyribonucleic acid,” abbreviated as “DNA.” Every single living cell contains such long “strings of letters.” And in multicellular organisms, they are exactly the same in every single cell of the body.
As for the information contained within, it can be thought of as codes within the variable sequences of A, T, G, and C. These codes are “read” by other cell organs as needed and then translated into various functions. The best known of these is the formation of variable proteins, i.e., protein building blocks. The physical substance of a living being consists of many different such proteins. In addition, the respective complete blueprint is also encoded in the genetic material. The complete code of a genome could therefore begin as follows and then continue with many more of the same four “letters” in varying combinations:
AGGATCCTACGTAGTGACCGTTGTCTAAGTCGACTTGATGTTGTCTAA (…)
This variable sequence of four identical “letters” contains, for example, codes for the external conversion of primary cells into the respective required protein-forming muscle cells, skin cells, liver cells, nerve cells, brain cells, or vitreous cells in mammals. And the “blueprint” also contained in the overall code ensures that, during the development driven by continuous cell division from the first fertilized egg cell to the finished animal with trillions of cells, the respective protein building blocks form in the right place. The cells at the site of the future eyes read the code for the vitreous body material according to the instructions in the overall plan and transform themselves externally into this material, so that the eyes can finally form in the fetus.
One can already imagine that such a genome—although the stored information is based on the same four “letters”—must be a truly incomprehensibly complex structure of intertwined codes. And with a little mathematics, this becomes even more impressive. Namely, when the number of possible combinations within the “letter series” is determined. The larger this number is, the higher the potential complexity of the stored information.
The formula used to calculate the so-called “variation with repetition” is suitable for mathematically determining these potentials within genomes. Written out in full, it is simply: (n^k). The number of types of nucleotide bases available, i.e., A, T, G, and C, becomes “n” (n=4), and the placeholder for the length of the sequence is “k.” This makes it quite easy to see how the result changes in relation to the possible number of combinations when the length of the genome strand, i.e., k, is gradually increased. Here are two examples of the results:
n = 4
k = 10
(4^10) = 1.048.576
n = 4
k = 200
(4^200)=
2.582.250.000.000.000.000.000.000.000.000.000.000.000.000.000.000.000.000.000.000.000.000.000.000.000.000.000.000.000.000.000.000.000.000.000.000.000.000.000.000
The result for k=10, with just over a million and thus seven digits, is not particularly spectacular. But obviously, increasing the series length k does not produce a linear result, but rather one that grows exponentially. Just by increasing it 20-fold to k=200, an incredibly gigantic number with 121 digits suddenly appears. In exponential notation, this may seem rather unspectacular again at 2.58 x 10^120. There are even names for such large numbers; in this case, it is called 2.58 vigintillion.
The estimated maximum number of all atoms in the universe pales in comparison to the number of possible combinations of even the smallest genomes
However, what this really means can be guessed by looking at what astrophysicists estimate to be the upper limit in another context: namely, the maximum possible number of all atoms in the universe. This actually refers to the totality of all atomic particles that make up the entire universe with all its galaxies and celestial bodies. Even the number of atoms in a single drop of water, at around five quadrillion (5^21), is beyond our imagination. The estimates for the entire universe range between 10^84 and 10^89 [3]. Even the highest estimate, 10^89, would fit around 10 quintillion times (factor 10^31) into the calculated number of 2.58 vigintillion possible variations at k=200.
But now something even more spectacular comes into play: there is no such “tiny” genome in the ecological structure of planet Earth with “only” 200 base pairs. The smallest genome strands of complete viruses are in the four-digit range. If we were to calculate the formula with k=1,000, the result would be so large that the estimated total number of atoms in the universe would be practically negligible in comparison. And for metabolizing organisms, such as bacteria or even plants and animals, the calculation becomes even more pointless. The smallest bacterial genome found to date was detected in the leafhopper endosymbiont Carsonella ruddii, with almost 160,000 sequences of A, T, G, and C. The result could not even be written out on this website.
Note: Regarding the virtually infinite potential complexity that genomes have as information storage and the resulting potential for variation in life forms, the properties of the chemical element carbon play a key role. Its atoms can form virtually unlimited variable and long chains with themselves and form many more compounds with other elements than all other known elements combined. These chemical fundamentals are further reflected in the section Universality.
Incidentally, it would be practically irrelevant if only a small proportion of the possible combinations of a genome were actually used for coding. So if only one-thousandth of the potential is used accordingly, then the absurdly long number is shortened by three digits.
The potential complexity of each genome, which extends almost to infinity and whose carrier exists in an ecological structure of unimaginably higher complexity, makes its selective breeding control from outside absolutely impossible – at least in the sense of a sustainable process.
Only if a life form aligns itself from generation to generation with its own primary benefit, automatically taking into account all the countless environmental influences – or rather, is automatically aligned by these influences according to its own benefit – can it remain stable in the long term. No matter how an external manipulator might try to permanently undermine this automatism in its favor, it could never succeed.
Supposed selective breeding patterns in nature are always based on misconceptions—this applies to all parasitic and symbiotic relationships as well as to the transmission of viral sequences
In the following, we will now examine some common misconceptions circulating in civilized society. One of these concerns the fact that genetic sequences previously developed in the evolution of viruses can be found in a variety of genomes across generations, for example in bacteria, plants, and animals. A typical misconception is the assumption that manipulation of traits across generations is taking place, since at least the change in the genome itself can be defined as a new trait that is passed on to subsequent generations of the host. There are two main reasons why this does not fulfill the breeding scheme for the primary benefit of a manipulator: First, these are not active viruses, but only fragments of the genetic material of former viruses. And second, the originally viral fragments integrated into the genetic material are neutral or useful for the organism, but not harmful.
The ancestors of the host organism were infected – mostly a very long time ago, several or many millions of years – and viral sequences that were useful to its lineage, for example because they were beneficial to the immune system, were retained and integrated almost automatically in the course of further evolution, as they brought a selective advantage. A significant proportion of the human genome – estimated at up to almost ten percent – also consists of viral sequences that have accumulated through integration into germline cells. They have established themselves as valuable or potentially useful components of the genome and often perform important functions, such as in the regulation of genes or the immune system.
Another misconception with some similarities revolves around certain bacteriophages, i.e., very small viruses that infect bacteria. Among these, there are some that, after infecting a bacterial cell, “decide” whether to replicate and destroy the bacterium – as typical viruses do – or instead remain neutral in the bacterial cell as a so-called “lysogenic” and thus non-harmful form. The lysogenic phage is then transferred to subsequent generations when the bacterium multiplies through cell division – not just in fragments, but as a complete virus. However, the bacteria are actually very interested in such an intake, because the presence of a lysogenic phage sequence gives them various advantages. One of these is a high level of immunity to further attacks by other phages that are potentially harmful rather than neutral. This immunity can also work against other “quasi-species” of phages and is then referred to as superinfection immunity. During the neutrality of the lysogenic phage, the sustainability parameter on the side of the bacterium is well above zero in the positive range, and the aforementioned pattern of disadvantageous breeding is not fulfilled.
Everything established so far also applies to the development of so-called “obligate symbioses.” Such a pattern exists when different species have adapted to each other to such an extent that they cannot survive independently without the other side(s). Well-known examples are lichens, which are obligatory symbioses between algae and bacteria that are so strongly intertwined that, when viewed from the outside, they appear to be a single organism.
Many of the bacteria living in our human digestive tract are also obligatory symbiotic partners of our body. We need them, for example, to break down nutrients, regulate acids, and defend against harmful microbes. However, these symbiotic partners never do any of this because our bodies have manipularly “bred” them to do so. Rather, over millions of years of evolutionary selection, they have automatically adapted to serve their own primary interests. Because our body is their habitat, their “home” so to speak, and – in the evolutionary sense of sustainability – is also supposed to be the home of their offspring, the development of characteristics that benefit us is something that primarily serves the interests of the bacteria.
From the evolutionary perspective of the microorganism, our benefit is secondary. Those siblings among the bacteria that provided the greatest possible benefit to their habitat always had a selective advantage and reproduced more frequently than those that were less successful or even harmed the host (i.e., their own habitat). As a result, the symbiosis grew to the mutual benefit of both parties.
Leafcutter ants do not cultivate fungi like crops—it is an ancient symbiosis between certain species
At first glance, obligatory symbioses sometimes appear to involve a “breeder” and a bred “useful organism” – in other words, a schematic similarity to our agricultural methods, in which the parameter of sustainability is well below zero on the manipulated side. However, these are always illusions. This is particularly common in literature on leafcutter ants and certain species of the fungus Armillaria mellea. Over the course of probably around 70 million years of evolution, these two life forms have adapted so strongly to each other that neither the ants nor the fungus could exist independently. And because the symbiosis works in such a way that the fungus, which has always lived mainly underground, is supplied with nutrients by the ants and in return produces small nodules that are ideal food for the ants, this is misinterpreted to mean that the ants have deliberately cultivated the fungus, just as today’s humans have done with numerous grains, fruits, and vegetables.
The obligatory symbiosis between leafcutter ants and the fungus Armillaria mellea is one of the relatively well-researched ecological relationships. There are thousands of studies and many scientists have invested a large part of their work in this area. However, not a single characteristic of the fungus or the ant has ever been proven to have been developed for the exclusive benefit of the other party and through manipulation of the same. The fungus was not “forced” by the ants to form nodules through breeding activities, but rather automatically “realized” through natural selection that this effort would benefit it because it would attract and bind the ants to itself.
Conversely, the ants were not selectively forced by the fungus to align themselves in such a way that they would supply it in the soil. Instead, each step of their own adaptations was always primarily for their own benefit from the evolutionary “perspective” of the symbiotic partners. The development of true obligate symbioses with pronounced physical characteristics is therefore a very long process, usually taking several or even many millions of years. And when the microorganisms involved are excluded, it is always only two or at most a few specific species that adapt to each other.
Not a single of agricultural varieties manipulated by humans is a mutualistic symbiotic partner
In order to recognize that obligate symbiosis in ecological structures can have only superficial similarities with human breeding practices involving “livestock” and “crops,” three clear differences can be identified (here, those involving plants and animals, excluding the microbes involved):
- In true obligate symbiosis, only a few particular species are very specifically aligned with each other. Usually, these are two or at most three species.
- The physical alignment of the characteristics of the partners relevant to the obligate symbiosis took place over periods of usually several million years.
- In obligate symbiosis, no characteristics can be found on either side that were forced upon the other side by targeted intergenerational manipulation for the primary benefit of the maniulator.
None of these points apply to any of the relationships with our agricultural “useful organisms.” We have not spent millions of years adapting specifically to one or two particular partner species without deliberately manipulating them across generations for our own primary benefit. Instead, within a few millennia, which is negligible in evolutionary terms, we have manipulated virtually every tangible life form across generations for our own benefit without restraint.
Thus, in no single case are the relationships between humans and the numerous cultivated “livestock” and “crops” a true obligate mutual symbiosis. One could try to classify them in the category of parasitic effects. However, this is also problematic because, as explained above, no parasitic life forms or viruses have ever been proven to exist in the Earth’s ecosystem that intervene in the evolution of their hosts in this way. And the reason, which has also already been discussed, is that this can never function sustainably due to the uncontrollable complexities involved and must always end in an evolutionary impasse.
Through selection for our primary benefit in terms of their own survival, “livestock” and “crops” have not gained any primary, and certainly no sustainable benefit or advantage. Instead, they have all become weaker in the face of countless and uncontrollably complex environmental influences.
Particularly in the final phase of intensifying this selection process for the purpose of creating more benefits for humans, the weaknesses have increased correspondingly.
It is sometimes argued that “farm animals” and “farm crops” have gained a significant advantage by being reproduced by humans and their populations being greatly expanded by numbers and geographically. However, this argument is invalidated when the parameter of sustainability is taken into account. For the majority of the plants and animals affected, its value is so far below zero that it can already be defined as ongoing extinction. The supposed “advantages” of the destabilization of their genotypes and phenotypes, as well as the inability of individuals to survive, which have arisen in tiny periods of time by evolutionary measures, are an illogical and absurd argument.
Today’s intensive agriculture marks the end of the evolutionary impasse
What we can now observe in the fields of monoculture or in the halls of the animal husbandry industry are organisms that are so weak in the face of the many components of environmental pressure that they have to be supported at great effort and become even weaker with each generation – a process that is further accelerated by increasing levels of manipulation. That is why they require increasing amounts of water, fertilizers, insecticides, herbicides, fungicides, antibiotics, antivirals, and many other aids.
The phase of intensification since the middle of the last century has led to a short-term sharp increase in the amount of food produced and, as a result, to an explosive increase in the human population from around 2.5 billion in 1950 to over eight billion in 2024. In the coming years of this dead-end situation, many different escalations will accelerate—if not slowed down—due to the inevitable further weakening of agricultural organisms and further consequential damage. Among other things, this will put increasing pressure on the ecosystem. It is already foreseeable that this unnatural and therefore unstable structure of civilized agriculture will inevitably collapse in the not too distant future. In particular, the application of newer genetic engineering techniques could act as a final powerful push in this fatal direction. Since the absolutely uncontrollable complexity is not understood, it cannot be understood that every increase in the degree of targeting to create one’s own benefit automatically pushes down the value of the sustainability parameter on the manipulated side.
The fact that the physical Law of Evolutionary Stability, despite its central importance and its factual confirmation by Charles Darwin, for example, is practically ignored in today’s natural sciences can be attributed to the same cause that also leads to the physical regularities discussed in the next section under the term “freedom” being largely ignored. These can also be explained in completely concrete and mechanical terms, even if the term might suggest something more philosophical. Furthermore, it will become apparent that the physical Law of Evolutionary Stability discussed here forms a solid foundation for the regular natural state of freedom. For since there is no sustainable possibility of selective breeding between different species, every living being of every species in the ecosystem is “born free” in the true sense of the word. A real, lasting relationship of subjugation between different species is therefore impossible in an ecological structure.
Conclusion
The Law of Evolutionary Stability (LES) has always determined life on Earth and is based on the high complexity of all genomes, which cannot be controlled from outside. It was the basis for the more than 500 million years of history of higher life on the planet and for its enormous diversity and complexity. Only in free evolution can such a system arise and remain stable. Moreover, this law of nature is the very foundation of the central regularity of freedom, because it excludes the mutual subjugation of different species. The very broad orientation of humans against these central laws and regularities of nature since the ‘Neolithic Revolution’ has led the system of civilisation into an evolutionary dead end. Numerous parallel escalations in the present suggest that its end is either imminent or has already been reached. It is of utmost importance to open up to these facts in order to find ways to defuse the situation.
[1] Charles Darwin, On the Origin of Species, 1859 Page 200 / 201. Source: http://darwin-online.org.uk/Variorum/1859/1859-200-dns.html
[2] Charles Darwin, On the Origin of Species, 1859 Page 29 / 30. Source: http://darwin-online.org.uk/Variorum/1859/1859-29-dns.html
[3] SWR Wissen: „Wieviele Atome gibt es im Universum“
https://www.swr.de/wissen/1000-antworten/wie-viele-atome-gibt-es-im-universum-100.html
[4] Written answer to Steffen Pichler on 05.08.2013 from the parasitologist Professor Dr. Richard Lucius, who has decades of research experience in various areas of his field and has also published specialised and non-fiction books providing a general overview of the range of parasitic strategies described. The question posed was whether he was aware of any parasitic strategies in which the parasite deliberately causes changes in the characteristics of the host’s subsequent generations for its own benefit. Professor Lucius’s answer:
‘That’s an interesting question, one that I’ve also asked myself. This could work, for example, through epigenetic changes induced by the parasite in the host’s germ line. However, only the manipulation of the current host using a wide variety of mechanisms is described, not that of its offspring.’